NDM-1—The Newest Recruit to the Army of β-lactamases
Sarah Turner, Ja’Nean Ray and Timothy Mietzner
In the recent popular press much discussion has been made of a new group of ‘super bugs’ that are emerging from India and Pakistan. These organisms have a specific resistance gene designated as NDM-1 (New Delhi metallo-β-lactamase-1). The gene encoding this β-lactamases associated with organisms that demonstrate multiple antimicrobial resistance profiles, limiting the treatment options for these infections.
NDM-1 was first described by Yong et al in December 2009 in a Swedish national who fell ill with an antibiotic-resistant urinary tract infection that he acquired in New Delhi, India. The infection was unsuccessfully treated in a New Delhi hospital which led to the patient's repatriation to Sweden. Klebsiella pneumoniae which expressed a novel metallo-β-lactamase (MBL) was identified as the source of the infection. This was the first report identifying the NDM-1 gene and this organism was susceptible only to fluoroquinolones and colistin. The authors concluded that the new resistance mechanism "clearly arose in India, but there are few data arising from India to suggest how widespread it is” (1).
In March 2010 the spread of NDM-1 was studied in a tertiary care unit in Mumbai, India. In 3 months, 22 different species of Enterobacteria were identified as NDM-1 producers. NDM-1 was so pervasive that only two Klebsiella pneumoniae carbapenem resistant strains did not produce the NDM-1 enzyme (2).
Much of the concern surrounding NDM-1 in the medical community is due to the fact that the gene has not been confined to a specific species of bacteria, which has largely been the case for methicillin-resistant Staphylococcus aureus (MRSA). Rather, this gene can be transferred to many different types of Enterobacteriaceae. Thus, this broad host range capacity gives it the potential to spread at an unprecedented rate (3).
The Indian response by members of Parliament disputed the claim made in the August 2010 publication of the Lancet that NDM-1 originated in India. The health ministry in India asserted that the alarm was spurred by Western doctors fearing loss of business to India by way of medical tourism. Prof. K.N. Prasad, a microbiologist at the Sanjay Gandhi Post Graduate Institute of Medical Science argued that the sensationalization over NDM-1 originating in India is unnecessary, saying “Already 40 types of bugs similar to NDM-1 exist globally, that includes the German imipenem-resistant metallurgic strain.
The Indian variant is just the 41st strain. These things happen.”[http://www.hindustantimes.com/Linking-India-to-superbug-unfair-and-wrong-says-India/Article1-585840.aspx ]. Professor Kumarasamy (University of Madras), the primary author of the 2010 Lancet study maintained that he does not agree with the part of the article that advises people to avoid elective surgeries in India. He said that many of the interpretations were added later without his permission or knowledge [http://timesofindia.indiatimes.com/city/chennai/Indian-author-says-superbug-report-is-fudged/articleshow/6302479.cms ]. NDM-1 isn't unique in attracting controversy over its name. From the colloquial term “Swine Flu” for H1N1 in 2009 to Syphilis being known as the “French disease” in 1530, denoting specific geographic locales as a name for a pathogen can be contentious.
β -Lactamases and NDM-1
β-lactamases are the primary causes of bacterial resistance to β-lactam antibiotics. These enzymes hydrolyze the β-lactam ring common to all penicillins and carbapenems (Figure 1). These enzymes are quite diverse in nature and can be organized into classes. In 1903, international efforts to construct the Panama Canal were being threatened by the twin biological hazards of yellow fever and malaria. The French decided to leave the endeavor in the hands of the Americans after enduring more than 12,000 deaths and an estimated $40 million loss. Tropical diseases were a deadly enemy, and if American efforts were to succeed), these diseases needed to be overcome.
Figure 1. Action of Beta Lactamase Cleaving the β-lactam
Ring.
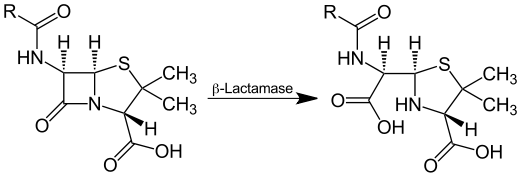
The two major classification schemes are the functional scheme proposed by Bush and Jacoby (4) and the molecular classification scheme proposed by Ambler (5). This Ambler classification is cited in the majority of publications concerning NDM-1. β-lactamases according to the Ambler system are classified by their amino acid sequence (Figure 2). Class A, C and D are different serine-β-lactamases. Class B refers to all the metallo-β-lactamases. There are similarities between the serine-β-lactamases that suggest they may have descended from a common ancestor. However, metallo-β-lactamases are very different from each other and probably are not related (6). Class A is the most diverse. The resistance can be encoded on plasmids, integrons, or on the bacterial chromosome (7).
Class A were the first β-lactamases described and hydrolyzed only simple penicillins. Later, resistance against both narrow and broad spectrum cephalosporins emerged and are now described as extended-spectrum β-lactamases (ESBLs) (8). A well known example of Class A ESBLs is found in the well described MRSA strains. This class also includes serine carbapenemases. Bacteria belonging to this group of β-lactamases are resistant to carbapenems, cephalosporins, penicillins, and aztreonam. Their multi-drug resistance is especially difficult to treat (7).
Class B contains all the metallo-β-lactamases including NDM-1. This class requires a bivalent metal ion, usually zinc, for the enzyme’s active site (6). Like Class A, the resistance in class B can be due to plasmids, integrons or chromosomes. Class B is usually resistant to everything except aztreonam. The plasmid carrying NDM-1 allows the bacteria to hydrolyze all β-lactams except for aztreonam (7). Because this plasmid is often transferred to bacteria that already has non-plasmid mediated resistance, it is able to hydrolyze all antibiotics except for tigecycline or colistin. In some cases, the bacteria is impervious even to these (3).
Figure 2.
Relationship of β-lactamases
as Described by Ambler.
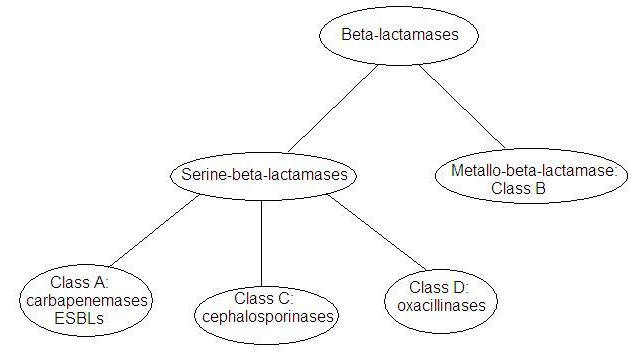
Class C β-lactamases were initially identified as being resistant to cephalosporins. The resistance is chromosomally mediated and is encoded by ampC (9). The transfer of ampC to bacteria through plasmids is becoming more prevalent. These bacteria are usually resistant to penicillins and cephalosporins. These bacteria poorly hydrolyze cefepime and are sensitive to cloxacillin, oxacillin, and aztreonam (7).
Class D are named oxacillinases because they were identified to hydrolyze isoxazolyl β -lactams such as oxacillin and methicillin. The resistance in this class is commonly conferred by plasmids and integrons (9). This class can usually hydrolyze penicillins, cephalosporins, extended-spectrum cephalosporins, and carbapenems. Currently, they are inhibited by known β-lactamase inhibitors clavulanate, sulbactam, and tazobactam with some exceptions (7).
Integrons and The Spread of Multiple Antibiotic Resistance
In 1986, the DNA sequence of several seemingly unrelated antibiotic resistance genes heralded the first hints regarding integrons (10). Common regions were noted upstream and downstream of various antibiotic resistance genes. These regions were found to be in different places on various plasmids, suggesting that, like transposons, these elements were mobile. However, the element differed from transposons in two important characteristics: (i) transposons have direct or indirect repeat sequences at their ends, but the regions surrounding the antibiotic resistance genes in the new elements were not repeats, and (ii) the elements contained a site-specific integrase gene of the same family as those found in phage but lacked many gene products associated with transposition. Due to these differences, the elements were not grouped with transposons and were named integrons.
Integrons are mobile DNA elements with the ability to capture genes, notably those encoding antibiotic resistance, by site-specific recombination. Integrons have an integrase gene (int), a nearby recombination site (attI), and a promoter. There are at least three classes of integrons based upon the type of integrase gene they possess. Class 1 integrons have been examined the most extensively. They consist of a variable region bordered by 5' and 3' conserved regions. The 5' region is made up of the int gene, attI, and the promoter which drives transcription of genes within the variable region. The 3' region consists of an ethidium bromide resistance locus (qacED1), a sulfonamide resistance gene (sulI), and an open reading frame containing a gene of unknown function. The integrase of Class 2 integrons is located within the 3' conserved region. Class 3 integrons have yet to be thoroughly studied. Examples of Class 1 integrons are shown in Figure 3.
Figure 3.
Examples of Class 1 Integrons
(taken from
http://www.sci.sdsu.edu/~smaloy/MicrobialGenetics/topics/transposons/integrons/integrons.html
)
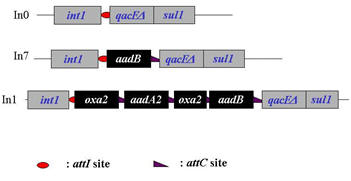
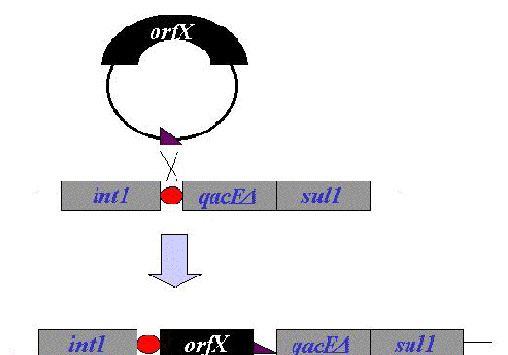
Figure 4.
Evolution of An Integron (taken
from
http://www.sci.sdsu.edu/~smaloy/MicrobialGenetics/topics/transposons/integrons/integrons.html)
In the original description
of the NDM-1 gene, Yong et al (1) prepared a genomic DNA
library from the resistant K. pneumoniae clinical isolate. This
library was transformed into a -lactam sensitive E. coli background and
selected for growth on β-lactam-containing medium. From this three distinct
antibiotic resistance conferring regions were identified (Figure 5). These
included genes for resistance to rifampin, erythromycin, gentamicin,
chloramphenicol, and three different β-lactamases (one of which is NDM-1). The
novel NDM-1 gene product displays tight binding to the penicillins and
cephalosporins. It is also active against the carbapenems. The only
antibiotics active in vitro against K. pneumoniae clinical
isolates were tigecycline and colistin. Perhaps more disturbing was the finding
of the NDM-1 gene on a 140-kb plasmid from a fecal E. coli isolate.
This indicates that this phenotype is highly transmissible and can be mobilized
to a broad host range.
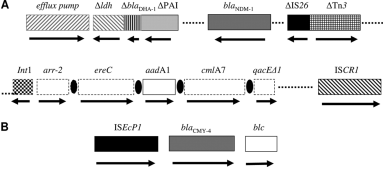
Figure 5. Three characterized antibiotic resistance-conferring regions
from K. pneumoniae 05-506. (A) The 4.3-kb region is linked to the
4.8-kb complex Class 1 integron region. The genes encoding the efflux pump and
lactate dehydrogenase (gray diagonal lines) are of Klebsiella origin. blaNDM-1
(dark gray) is flanked between the pathogenicity island (vertical black lines)
and IS26/Tn3 (black small squares). This region lies downstream of the 4.8-kb
complex Class 1 integron containing Int (checkered area), arr-2, ere2A, aadA1,
and cmlA7 as gene cassettes and qac 1 (white boxes). Downstream is an intact
copy of ISCR1 (black and white diagonal lines). Arrows, direction of
transcription; black ellipses, 59-base elements;
genes that are truncated. (B) blaCMY-4 (gray) is located
between ISEcP1 (black) and blc (white).
Epidemiology of NDM-1
As of November 2010 reports of NDM-1 producers were identified in India, Pakistan, the UK, the Netherlands, Japan, Australia, France, Spain, Sweden, Finland, Denmark, Austria, Belgium, Norway, Italy, Oman, Kenya, and the Balkan region. The majority of these patients had recently traveled to the Indian subcontinent.
In November 2010
seventy-seven patient cases were reviewed in European countries. Among these
cases, four patients developed a blood-stream infection. Seven died in the
hospital and one was directly related to NDM-1 positive E. coli which
caused septic shock in a leg wound. Most of these patients had recently (within
the last year) traveled to the Indian subcontinent or the Balkan region, but
other areas included Dubai and Spain. The majority of these patients had been
admitted to the hospital because of illness or an accident, and a small minority
of hospitalizations were due to medical tourism (6).
Two patient cases in Italy had no travel history, but were in a hospital unit where a patient returning from India had stayed. Another two patients in the UK had no travel history and could not be linked to another travel-associated case. In some isolates in this study patients tested positive for the NDM-1 enzyme in K. pneumoniae from urine and E. coli from feces suggesting in vivo transfer. At this moment evidence of secondary nosocomial transmission of the NDM-1 enzyme in Europe is limited.
The patients who are most susceptible to a severe illness due to bacteria expressing NDM-1 are those with comorbidities and/or needed invasive care procedures (6). In the Indian subcontinent a pertinent concern is the NDM-1 is commonly found in E. coli which is the number one cause of diarrhea in children. Antibiotic resistance in this population can have potentially devastating consequences (3). Indirect fecal-oral transmission and exposure to contaminated people, food and water are the most likely ways these pathogens are spread. Countries that lack an adequate sanitation infrastructure are in the greatest danger of having NDM-1 community based infections. India is identified as a reservoir for NDM-1 caused by an enormous population of people living in these conditions (11). It is possible that other areas lacking this infrastructure are at risk of becoming NDM-1 endemic. Patients hospitalized in the Balkan region were positive for NDM-1 producers. Between 2007- 2009 NDM-1 positive K. pneumoniae was found in patients in a tertiary care center in Nairobi, Kenya. These patients had no travel history to the Indian subcontinent or the UK, but there is a large population of Indian diaspora to Kenya (12, 13).
Clinicians and scientists
are advocating preventative measures of NDM-1 infections since pharmaceutical
options are severely limited. Overuse of the few effective antibiotics poses a
risk of antibiotic resistance to those medications.
NDM-1, like other MBL’s (metallo-β-lactamases), hydrolyzes all β-lactam antibiotics except for aztreonam but it is usually inactivated by co-produced extended-spectrum β-lactamases. Most of the bacteria producing the NDM-1 enzyme belong to the Enterobacteriacea family and are usually only susceptible to colistin and occasionally tigecycline (14).
The CDC recommends using the guidance outlined for carbapenem-resistant Enterobacteriaceae for treating possible cases of NDM-1. This guideline includes identifying the pathogen as carbapenem-resistant and placing patient into contact precautions. Some circumstances may call for point-prevalence surveys or surveillance testing among other high-risk patients.
The CDC is asking that
carbapenem-resistant isolates from patients who have received medical care in
India or Pakistan for the last six months be forwarded through the state public
health laboratories to the CDC to determine whether or not the bacteria is NDM-1
producing. Infection control by preventing transmission is of the utmost
importance when managing an NDM-1 positive patient (http://www.cdc.gov/mmwr/preview/mmwrhtml/mm5924a5.htm).
1. Yong D, Toleman MA, Giske CG, Cho HS, Sundman K, Lee K, Walsh TR. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother, 2009;53:5046-54. [PubMed]
2. Deshpande P, Rodrigues C, Shetty A, Kapadia F, Hedge A, Soman R. New Delhi Metallo-beta lactamase (NDM-1) in Enterobacteriaceae: treatment options with carbapenems compromised. J Assoc Physicians India, 2010;58:147-9. [PubMed]
3. Nordmann P, Poirel L, Carrër A, Toleman MA, Walsh TR. How to detect NDM-1 producers. J Clin Microbiol. 2011;49(2):718-21 [PubMed]
4. Bush K, Jacoby GA, Medeiros AA. A functional classification scheme for beta-lactamases and its correlation with molecular structure. Antimicrob Agents Chemother, 1995;39(6):1211-33. [PubMed]
5. Ambler RP. The structure of beta-lactamases. Philos Trans R Soc Lond B Biol Sci, 1980;289:321-31. [PubMed]
6. Garau G, Di Guilmi AM, Hall BG. Structure-based phylogeny of the metallo-beta-lactamases. Antimicrob Agents Chemother, 2005;49:2778-84. [PubMed]
7. Drawz SM, Bonomo RA. Three decades of beta-lactamase inhibitors. Clin Microbiol Rev 2010;23:160-201. [PubMed]
8. Coque TM, Baquero F, Canton R. Increasing prevalence of ESBL-producing Enterobacteriaceae in Europe. Euro Surveill 2008;3:9. [PubMed]
9. Kong KF, Aguila A, Schneper L, Mathee K. Pseudomonas aeruginosa beta-lactamase induction requires two permeases, AmpG and AmpP. BMC Microbiol 2010;10: 328. [PubMed]
10. Cambray G, Guerout AM, Mazel D. Integrons. Annu Rev Genet, 2010;44:141-66. [PubMed]
11. Webster PC. Alarm grows over management of antibiotic resistance file. CMAJ 2010;182(2):E141-2. [PubMed]
12. Poirel L, Al Maskari Z, Al Rashdi F, Bernabeu S, Nordmann P. NDM-1-producing Klebsiella pneumoniae isolated in the Sultanate of Oman. J Antimicrob Chemother 2011;66:304-6. [PubMed]
13. Poirel L, Revathi G, Bernabeu S, Nordmann P. Detection of NDM-1-Producing Klebsiella pneumoniae in Kenya. Antimicrob Agents Chemother, 2011;55(2):934-6. [PubMed]
14. Struelens
MJ,
Monnet DL,
Magiorakos AP,
Santos O'Connor F,
Giesecke J;
European NDM-1 Survey Participants. New Delhi
metallo-beta-lactamase 1-producing Enterobacteriaceae: emergence and response in
Europe. Euro Surveill, 2010;15(46),pii: 19716.